Аннотация:
В данной статье приведен обзор, классификация и сравнительный анализ гипотез эволюционного происхождения рода Unicornis на основе данных современной систематики надотряда Paraxonia без учета влияния магических полей на филогенез таксона. Также приведены основные аргументы в поддержку или против различных взглядов на эволюцию Unicornis и указаны наиболее актуальные в научном сообществе мнения касательно вопроса классификации данного рода.
Abstract:
This article provides an overview, classification, and comparative analysis of hypotheses of the genus Unicornis evolutionary origin based on the data of the superorder Paraxonia modern taxonomy without taking into account the influence of magic fields on the the taxon phylogenesis. The main arguments in support or against different views on the evolution of Unicornis are also given and the most relevant opinions in the scientific community regarding the classification of this genus are indicated.
Ключевые слова: единороги, копытные, китообразные, мезонихии, эволюция и систематика парнокопытных, Artiodactyla, Mesonychia, Cetancodontamorpha, Unicornis
Keywords : unicorns, ungulates, cetaceans, mesonychia, evolution and taxonomy of artiodactyls, Artiodactyla, Mesonychia, Cetancodontamorpha, Unicornis
DOI: f7e5eff3efe5ebe0
УДК: e6e0e1e0
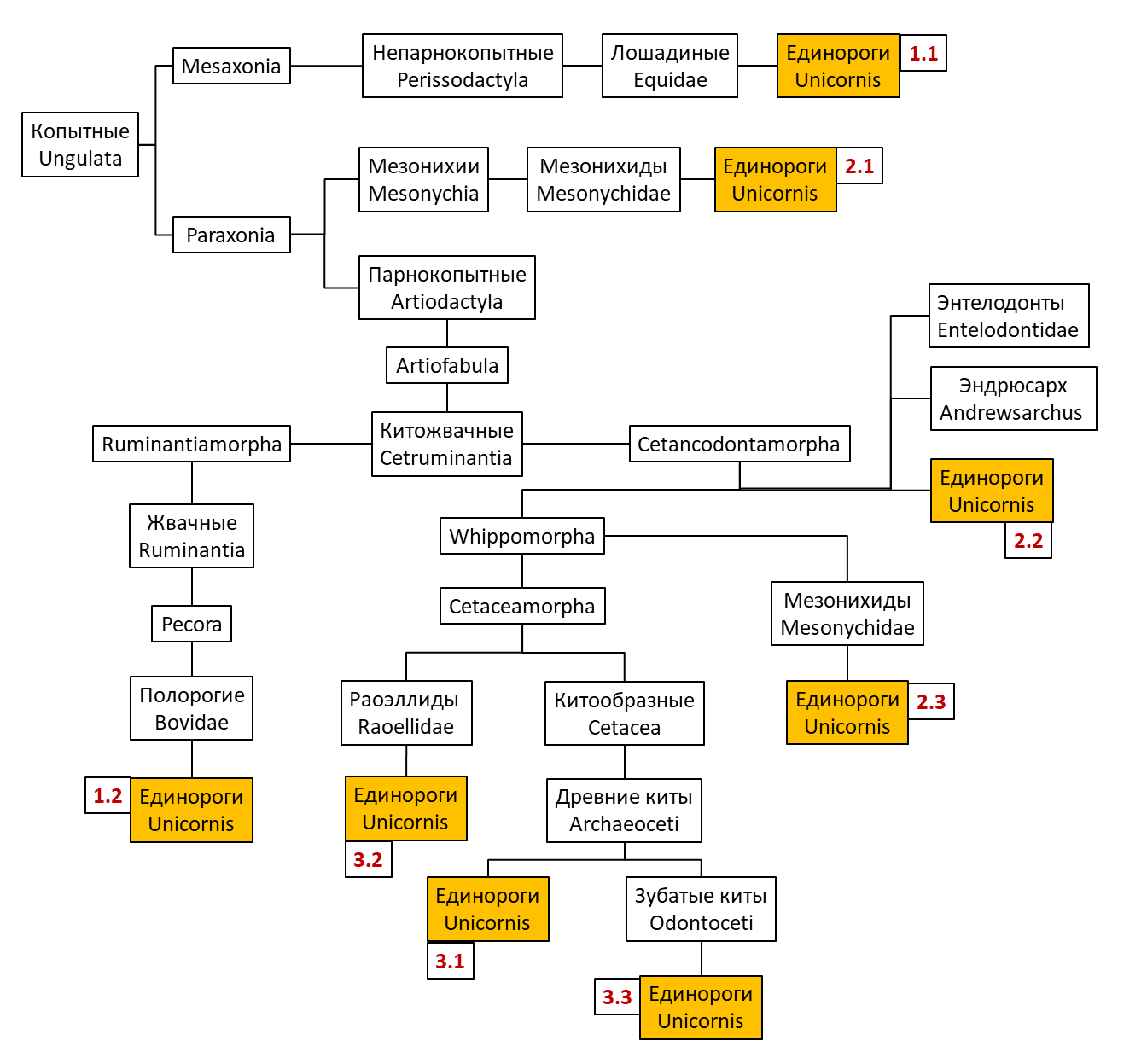
Рис. 1. Обобщающая схема гипотез происхождения рода Unicornis. Схема может не отражать истинной эволюционной и систематической иерархии таксонов и служит исключительно для наглядного представления материала.

Рис. 2. Сравнение внешнего вида Equus ferus caballus (а) и Unicornis Mirabilis (б).

Рис. 3. Нормальные (а,б) и “искаженные” (в-ж) формы рогов Unicornis Mirabilis: а — прямой, б — серповидный, в — искривленный, г, д — ветвистый (число ветвей не может быть больше 2), е, ж — двойной. Все “искаженные” формы наблюдаются крайне редко и являются следствием травмы, болезни, либо врожденной мутации.

Рис. 4. Сравнение малых коренных зубов Mesonychia на примере мезоникса (Mesonyx sp.) (а) и Unicornis Mirabilis (б).

Рис. 5. Andrewsarchus mongoliensis, реконструкция.

Рис. 6. Реконструкции представителей групп, близких к возможным предковым формам рода Unicornis: а — Pakicetus inachus, б — Indohyus major.
Аргументами в ее пользу являются особенности строения зубов U. Mirabilis, а также их (предполагаемый) изначально семиакватический образ жизни. Однако, большая часть ранних Cetacea ассоциирована с теплыми водами, и пока что нет достоверных сведений о массовой миграции подобных животных (Клеверова, 2003). При этом, если считать предковой формой полуводные виды, то их сухопутная миграция на север и вовсе не представляется возможной. И, хотя на данный момент представители рода Unicornis широко распространены по всему миру, палеонтологическая летопись свидетельствует о том, что их родиной являются северные приполярные области, а именно приморская тундра и, в прошлом, тундростепь (Золотов, 1916, Несси, 2000).
Гипотеза 3.2 определяет единорогов не в группу китообразных, но вместо этого сближает их с переходными формами, более приспособленными к жизни на суше, в частности с семейством Raoellidae (Рогов, 2017) (рис 6, б). На данный момент эта гипотеза считается наиболее актуальной, так как объясняет как сходство U. Mirabilis с продвинутыми Artiodactyla, так и особенности их образа жизни и питания. Raoellidae уже вели околоводный образ жизни, но при этом обладали развитыми конечностями с копытами и могли мигрировать на север вдоль крупных рек, приспосабливаясь к новым условиям и, в конце концов, дав начало предковой форме U. Mirabilis. Пока что нет палеонтологических данных, что подтверждали бы эту гипотезу, однако можно предположить, что предок U. Mirabilis представлял собой некрупное, всеядное, рогатое парнокопытное (вероятнее всего, еще двурогое, но уже с тенденцией сращивания рогов), обитавшее по берегам северных рек и морей. Скорее всего, на этом этапе вид еще не обладал магическими способностями, но уже должен был начать испытывать на себе преобразующее действие магических полей (Кракелюров, 2000).
Гипотеза 3.3 считается самой экстравагантной из списка и зачастую не учитывается научным сообществом, однако, за свою оригинальность, заслуживает упоминания. Согласно ей, U. Mirabilis относятся относятся к зубатым китам (Odontoceti), которые вторично вышли на сушу уже после того, как заселили океаны и таким образом достигли северных земель (Воронов-Вальдфогель, 2016). И, хотя подобное событие объяснило бы появление предков единорогов на севере при отсутствии переходных форм между Raoellidae и современными единорогами, вероятность вторичного выхода на сушу водных животных представляется маловероятной. Сила же магического воздействия, понадобившегося для подобного преобразования, недостижима в естественных условиях (Кракелюров, 2000).
Заключение
Подводя итоги, можно заключить, что, хотя бытовой подход к классификации U. Mirabilis и повлиял на традиционную терминологию, он не может считаться хоть сколько-нибудь научным.
Что касается более поздних гипотез, то наиболее актуальна на данный момент гипотеза происхождения U. Mirabilis от семейства Raoellidae или сходных с ним форм околоводных животных, являющихся переходной формой между Cetacea и другими Artiodactyla. Происхождение от более развитых Cetacea помешало бы сухопутной миграции. Однако на данный момент пока еще не найдено палеонтологических подтверждений данной гипотезы.
Несмотря на пересмотр положения и статуса Mesonychidae в системе природы, нельзя исключать вероятность происхождения U. Mirabilis от родственных семейству Mesonychidae форм, в таком случае приспособление к околоводному образу жизни у предков U. Mirabilis произошло уже после того, как они мигрировали на север. Впрочем, и в этом случае только предстоит отыскать палеонтологические подтверждения, которые смогли бы поставить точку в вопросе эволюции и таксономического положения единорогов.